晩秋の野山の岩陰や家屋の片隅にナミテントウ(Harmonia axyridis Pallus、ナミ)が集まっている写真をみかけることがあります(谷岸、1976、高橋、1995)。ナミが冬を越す準備をしているのです。

写真1 ナミ越冬集団の捕獲場所
ガスボンベを中心にして、面積4m2の範囲以内にナミ成虫は飛来、壁に留まり、屋根に向かって歩いて、建物の中に消える。
昆虫は、冬が近づくと、あまりエネルギーを使わない卵や蛹の形態となって冬を過ごします。冬の間は餌となる動植物もなく、寒さも厳しいので自らの代謝機能を春の餌のある時期と上手く揃うように調節しているのです。

写真2 ナミ群団
二階西側の窓枠に集まる。
ナミ成虫が集団で越冬している写真をみると、「何故、成虫態で越冬するのだろうか」「集団で越冬することにどんな意味があるのだろうか」「冬の前まではバラバラに生活していたのにどのようにして集合するのだろうか」などと考えてしまう。
これらの疑問に対しては、研究者仲間でもいろいろな意見があります。虫たちが集団で越冬すれば、1)天敵などの攻撃に対して捕食の危険を減少させる防衛効果があるからだ、2)集団生活は単独よりも乾燥などの生息環境をより良い状態に保つことができ、生存率を高めることができるからだなどの考え方があるのです(近ら、2003)。
しかし、これらの考え方については納得できる説明がされていません。冬の間ナミ集団が天敵に襲われている写真もなければ、生息密度効果が生存率に及ぼす実証実験も行われていません。後者の場合は、高密度環境下では何故生存率が高いのかの科学的な証明がなされていないのです。
ある日、つくば市郊外の自宅の白壁にナミ群団が飛来する姿を目撃した(写真1&2)。ナミの野外観察を始めて、多少の知識をもっていたが、初めての経験であったので驚いた。研究者“たましい”が再びめばえて、ナミを捕獲して個体数や斑紋型分布比率を調べた。すると、これまで報告されているナミの斑紋型の変異に関するデータと異なっていた(駒井ら、1956)。そこで、数年間の観測結果をまとめて発表した(木村、2008)。
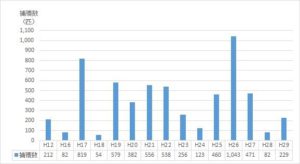
図1 年次別捕獲数の変動
しかしながら、ナミの捕獲数と斑紋型分布比率との関係は多少理解されたものの、越冬場所への飛来と気象などとの関係については一定の法則性をみだすことができなかった。科学的なデータを得るためには、さらなる観測が必要であったのだ。
今回は、これまでの観測データに新しいデータを加えて、ナミの越冬飛翔行動を詳細に解析、繁殖・生存戦略について考察した。
ナミの越冬飛来集団の年次別変動
ナミの集団飛来は平成12年に確認され、その後平成16年から平成29年までの14年間に亘って毎年捕獲個体数を調べた(図1)。15年間の捕獲数は5886匹で、平均393匹であった。最高の捕獲数は、平成26年の1043匹、最低は平成18年の54匹であった。捕獲数のばらつきを標準偏差で求めると、平均値±標準偏差値は392±277匹であった。このことから、捕獲数、すなわち飛来数は年度によって大きく異なることが明らかになった。
次いで、各年度の越冬飛来集団の飛来(捕獲)回数と捕獲数を調べた(図2)。最初の集団が現れる日は年次によって異なり、最も早いときは平成21年の11/9で、最も遅い日は平成24年の12/6日であった。飛来が一度ののみ大量にみられた年次は平成16年、18年、28年などの3回で、連続的にみられた年次は平成17年、19年、20年、21年、22年、23年、24年、25年、26年、27年、29年などの11回であった。それらの事象を一覧表にまとめた(表2)。
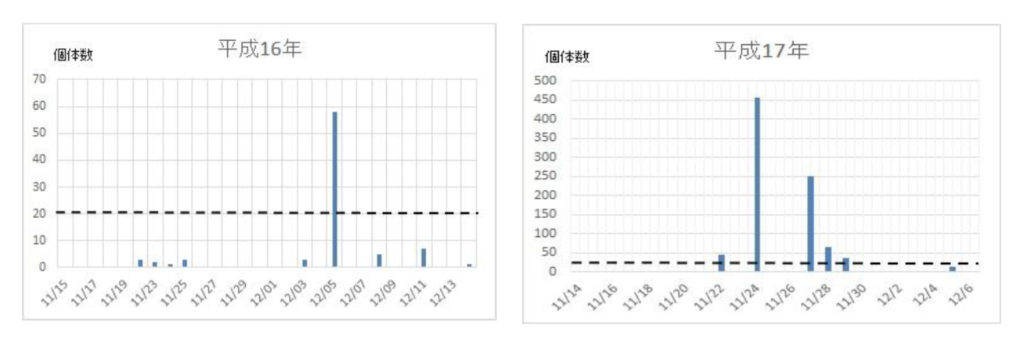
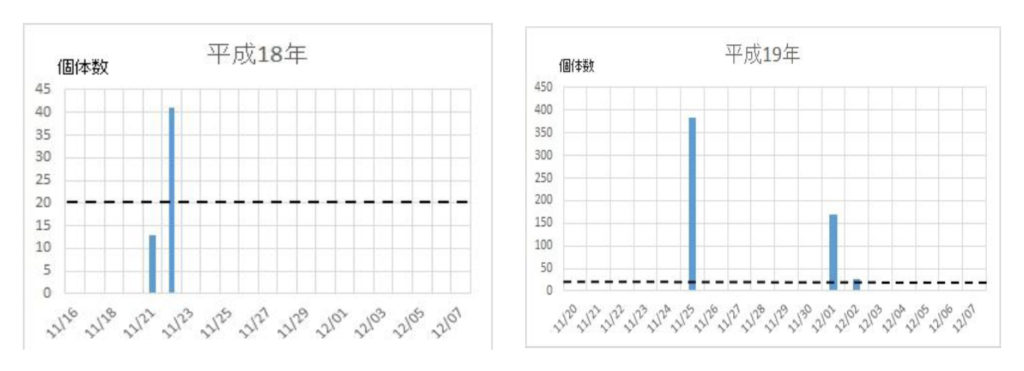
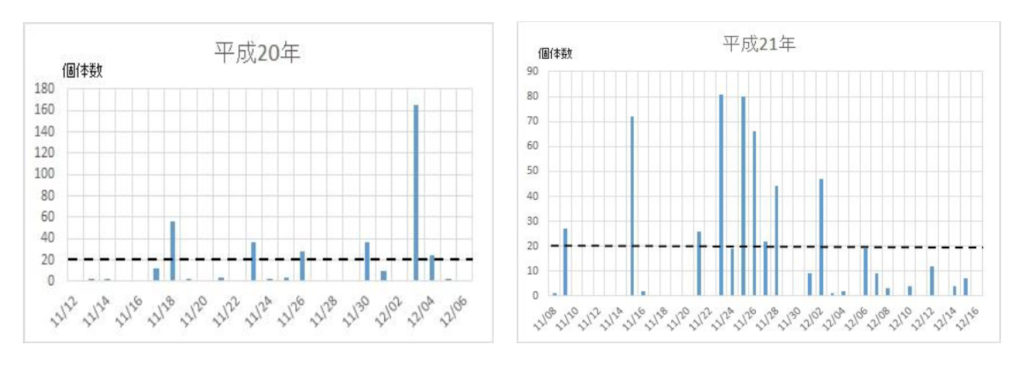
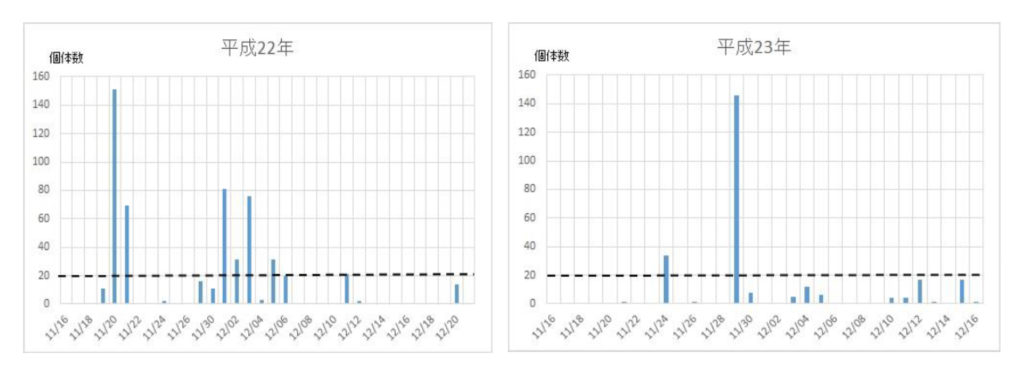
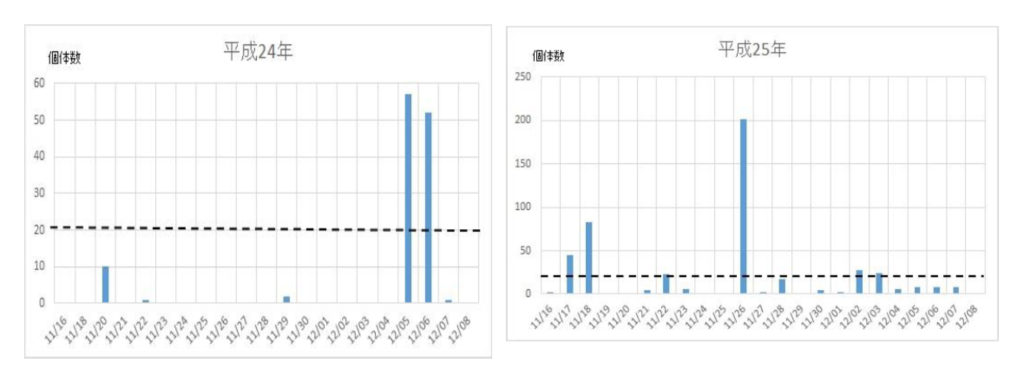
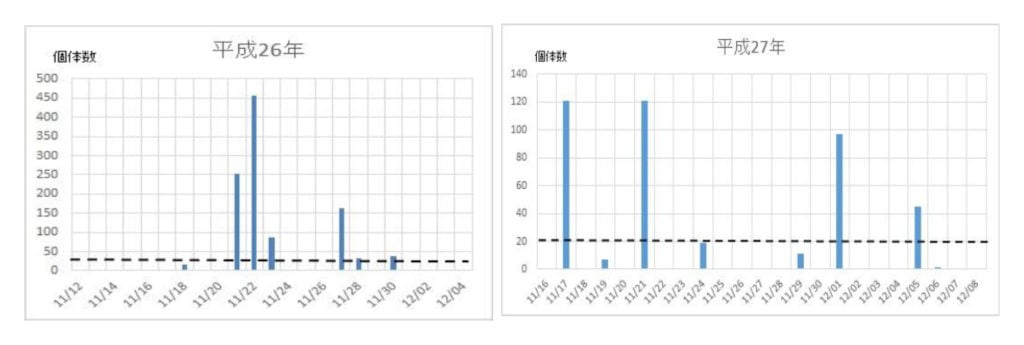
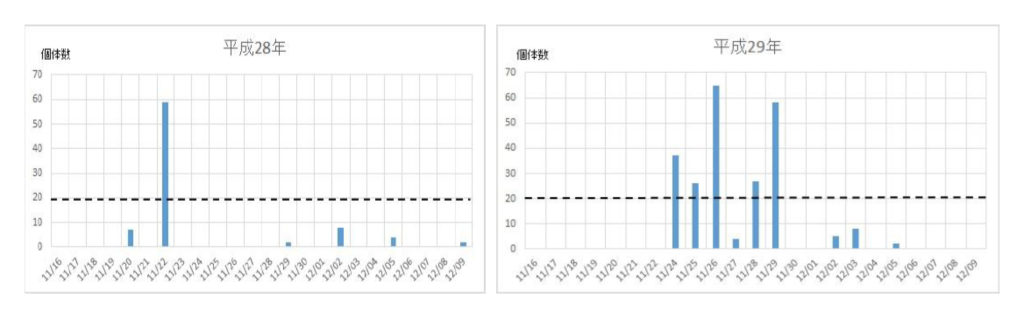
図2 ナミテントウの各年次における捕獲日時と捕獲数の関係
ナミの飛来は、11/21~11/25の5日間を中心として起こることが分かった(表1)。また、飛来が数十日に亘って続く場合と1日のみの場合があったので、前者の場合は複数の繁殖地から飛来し、後者の場合は一カ所の繁殖地からであると考えられた。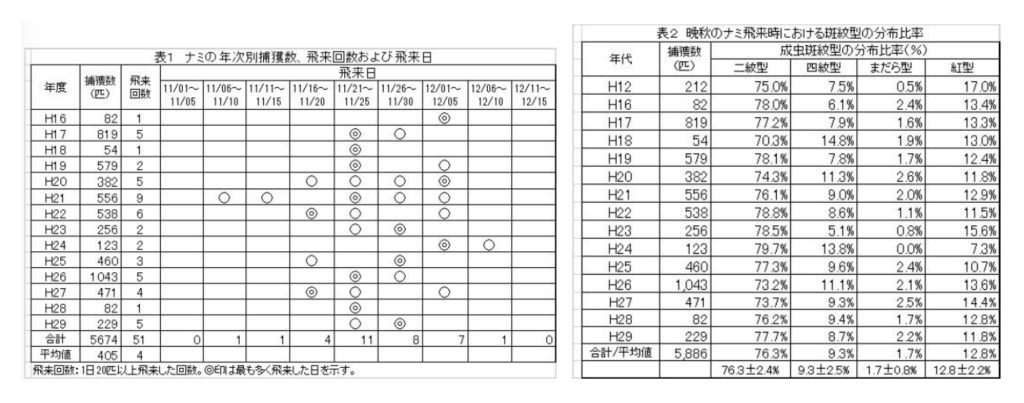
これらのことから、ナミの越冬飛来集団の飛来数と飛来回数は、繁殖集団の個体群の大きさと個体間の発育の差に依存し、繁殖特性によると思われた。
日別飛来数と気象条件および斑紋型分布比率の関係
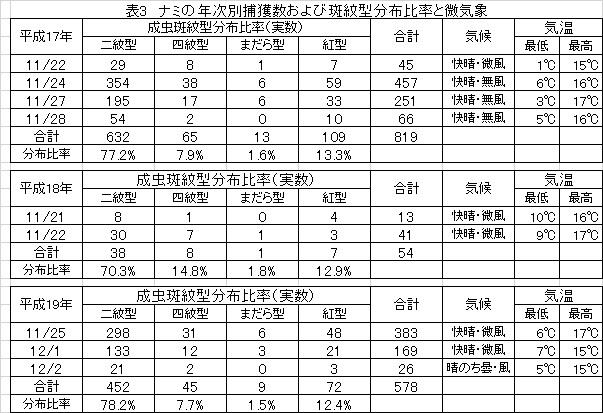 ナミの飛来行動を促す要因を知るため平成17年、18年、19年の3年間のナミ捕獲時の天候および斑紋型分布比率の変動を調べた(表3)。
ナミの飛来行動を促す要因を知るため平成17年、18年、19年の3年間のナミ捕獲時の天候および斑紋型分布比率の変動を調べた(表3)。
ナミの飛来時の気候は快晴・無風であるが、雲が出て陽が陰り、風が吹くとナミは途端に飛んで来なくなった(表3:平成19-12/2)。また、この時の気温は最低5℃前後、最高15℃前後であった。この寒暖の温度差によって、落ち葉などの下にいたナミは家屋の白壁から発する熱線に誘引されて飛翔行動を起こすものと考えられた。
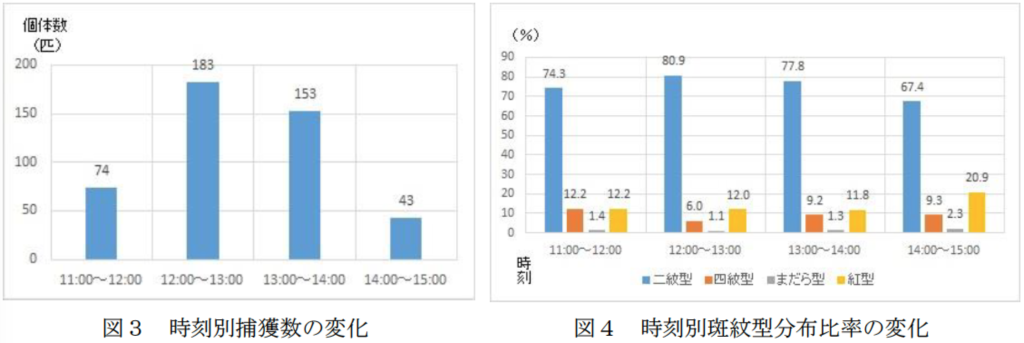
時刻別斑紋型分布比率の変動
ナミが遠赤外線に誘引されて越冬飛翔するならば、二紋型などの黒化型と紅型の非黒化型間の紋様の違いが飛翔行動に影響を与え、分布比率に違いが表われるのだろうか。そこで、平成17年11/24の大飛来集団の時刻別飛翔数(捕獲数)と斑紋型の分布比率を調べた(図3および4)。1時間あたりの捕獲数をみると、正規分布を示し、ひとつの山形であった。これは、ナミは日射量の多少によって飛翔すると思われた。とくに、午後3時以降はまったく飛来が見られなかったことは、白壁から反射する遠赤外線の減少によると思われた。一方、時刻別の斑紋型分離比率はほぼ一定で、変化しなかった。これは、黒化型と非黒化型も熱線に対しては同じように反応したということを意味する。そこで、メラニン系色素によってエネルギーを吸収して活発な行動を促すというthermal melanism 説は否定された。このことは、むしろ、遠赤外線受容器をもつある種の甲虫類の走熱性メカニズムと類似するものと思われた。